30秒注册登陆,可查看更多信息,结交更多好友,享用更多功能,轻松玩转论坛,白白手拉手欢迎您的加入!
您需要 登录 才可以下载或查看,没有账号?注册新成员
x
抗原特异性通过 Tregs 增强白癜风的疾病控制
Zhussipbek Mukhatayev 1,2,3,4* , Emilia R. Dellacecca 1,2 , CormacCosgrove 1,2 , Rohan Shivde 1,2 , Dinesh Jaishankar 1,2 , Katherine Pontarolo-Maag 5 , Jonathan M. Eby 5 , Steven W.Henning 5,Yekaterina O. Ostapchuk 4,Kettil Cedercreutz 1,Alpamys Issanov 6,Shikhar Mehrotra 7,AndreasOverbeck 8,Richard P.Junghans 9,Joseph R.Leventhal 10和I. Caroline Le Poole 1,2*
- 1美国西北大学皮肤科,芝加哥,伊利诺伊州
- 2 Robert H. Lurie 综合癌症中心,西北大学,芝加哥,伊利诺伊州,美国
- 3哈萨克斯坦阿拉木图国立大学生物与生物技术学院
- 4分子免疫学和免疫生物技术实验室,MA Aitkhozhin 分子生物学和生物化学研究所,哈萨克斯坦阿拉木图
- 5美国伊利诺伊州梅伍德洛约拉大学肿瘤研究所
- 6哈萨克斯坦努尔苏丹纳扎尔巴耶夫大学医学院医学系
- 7南卡罗来纳医科大学外科,南卡罗来纳州查尔斯顿,美国
- 8色素障碍外科,Lumiderm,马德里,西班牙
- 9美国波士顿大学血液学/肿瘤学系
- 10综合移植中心,西北纪念医院,芝加哥,伊利诺伊州,美国
白癜风是一种以黑色素细胞破坏为特征的自身免疫性皮肤病。白癜风皮肤中的调节性 T 细胞 (Tregs) 大大减少,补充外周皮肤 Tregs 可以防止色素脱失。神经节苷脂 D3 (GD3) 被包括黑素细胞在内的病灶周围表皮细胞过度表达,这促使我们产生 GD3 反应性嵌合抗原受体 (CAR) Tregs 来治疗白癜风。小鼠接受未转导的 Tregs 或 GD3 特异性Tregs 以检验抗原特异性有助于降低体外和体内自身免疫反应性的假设. 在自发性白癜风的 TCR 转基因小鼠模型中,与未转导的 Tregs 和载体对照受体相比,CAR Tregs 显示出响应抗原的 IL-10 分泌增加,对黑素细胞的细胞毒性提供了更好的控制,并支持显着延迟脱色。与两个对照组相比,后一项发现与治疗小鼠中更多丰富的 Tregs 和黑素细胞有关。我们的数据支持可以制备、使用和储存抗原特异性 Treg 以长期控制进行性脱色的概念。
介绍
白癜风是一种自身免疫性疾病,其中黑色素细胞被逐渐破坏,导致皮肤苍白、白色斑块 ( 1 , 2 )。白癜风的患病率为0.5-1 %,这种疾病与自尊心低下、心理压力和社会耻辱感有关 ( 3-5 )。目前可用于白癜风的疗法疗效有限,并且确实需要新的治疗策略 ( 6 )。有几个因素促成了白癜风的发病机制。黑素细胞中的氧化应激可触发诱导型热休克蛋白 70 (HSP70i) 的释放 ( 7)。HSP70i 可以直接激活先天免疫细胞和伴侣黑素细胞抗原,以便随后引入抗原呈递细胞 (APC) ( 8 , 9 )。反过来,这些先天免疫细胞和 APC 会募集自身反应性 T 细胞来介导白癜风中黑素细胞的特异性破坏 ( 10 )。对自身抗原的耐受性丧失还涉及皮肤调节性 T 细胞 (Treg) 活性降低( 11 , 12 )。Tregs 是 T 细胞亚群,负责通过抑制免疫细胞(包括自我反应性、细胞毒性 T 细胞)来维持免疫稳态(13)。在几种自身免疫性疾病中报告了Tregs 的缺乏和缺陷 ( 14 – 16 )。在病灶、非病灶和病灶周围部分的白癜风皮肤中发现了局部 Tregs 缺乏,从而表明白癜风患者维持外周免疫稳态的能力降低 ( 17 , 18 )。其他人报告说,Tregs的循环数量可能减少或表现出降低的活性 ( 16 , 19 )。白癜风皮肤中有限的Treg 活性允许自身反应性、细胞毒性 T 细胞扩增并消除皮肤中的黑色素细胞,从而促进进行性色素脱失 ( 12 )。
迄今为止,多克隆 Tregs 的过继转移已用于几种情况的临床试验,其中自体细胞在体外扩增并重新给予患者,但成功有限 ( 20 )。在白癜风的小鼠模型中,这种方法是有益的,通过在这种情况下短期维持有利的 Treg 与效应 T 细胞比率( 21 )。然而,多克隆Tregs 的应用可以通过无意中抑制对感染或恶性肿瘤的反应而产生全身免疫抑制,这是一个重要的临床考虑(22)。此外,产生足够数量的用于临床的多克隆 Treg 可能具有挑战性。通用免疫抑制可以提出一个问题,可以通过包括对过继转移细胞的抗原特异性来克服这一问题。在移植生物学中,抗原特异性 Treg 有望通过靶向供体和受体之间的人类白细胞抗原 (HLA) 差异在疾病进展部位提供局部免疫抑制 ( 23 )。抗原特异性Treg 也可能为白癜风的治疗提供显着优势。
抗原特异性 Treg 可以通过引入 T 细胞受体 (TCR) 来产生,但这些 TCR-Treg 的应用受到主要组织相容性复合体 (MHC) 限制的限制,无法让所有患者受益 ( 24 )。相反,我们的研究利用转导的抗原特异性 Tregs 来表达嵌合抗原受体 (CAR),从而克服了 MHC 依赖性。在 CAR-T 细胞治疗中,优化和选择正确的 CAR 亲和力和细胞内信号结构域对于 Treg 的治疗活性和细胞持久性尤为重要。由于 CAR 是使用抗体可变区构建的,因此与 TCR 相比,它们对其同源抗原具有更高的亲和力(25)。同样重要的是识别在研究中受影响的组织中相对丰富的细胞表面抗原,并且可以被 CAR Tregs 靶向。这个概念是,选择的表面抗原可以作为抗原特异性 CARTregs 的靶标,以选择性地和局部地抑制白癜风中的自身反应性 T 细胞 ( 26 )。这些结果可以与使用酪氨酸酶反应性 TCR 获得的结果进行比较。后一种方法在免疫缺陷黑色素瘤模型中进行了测试,首次证明了对过继转移的抗原特异性 Tregs 的活性 ( 27 )。酪氨酸酶和HLA-A2 编码基因都与白癜风疾病的发展相关,从而与酪氨酸酶反应性 T 细胞以及相关的 HLA-A*0201 限制相关。28 ). TCR 和 MHC 转基因的相同组合用于生成白癜风的 h3TA2 小鼠模型,这里用于宿主我们的 CAR 转基因 Tregs ( 29 )。这些小鼠出现快速、持续的色素脱失,大多数分化的色素细胞在 5 周龄时从皮肤上消失(30)。这种侵袭性的疾病模型为治疗的有效性设定了很高的标准,并且可以为新的治疗提供疗效的初步估计 ( 30 )。与此同时,神经节苷脂 D3 (GD3) 抗原具有相关性,因为 GD3 表达存在于受压的黑素细胞中,并且在白癜风患者的病灶周围皮肤中可能更为显着 ( 31)。这种 GD3 抗原目前被靶向用于黑色素瘤的免疫治疗,因为它在膜上的表达和抗体的可及性 ( 32 , 33 )。此外,GD3 有助于黑色素生成、细胞生长和细胞树突状 ( 34 , 35 )。为了测试抗原特异性Tregs 治疗白癜风的治疗潜力,我们分离了幼稚 T 细胞并将它们极化为 Tregs,然后引入 GD3 响应性CAR 编码构建体。在体外测试产生的细胞对相关靶标的细胞因子分泌,并测量产生的细胞毒性。最后,在进行性白癜风的小鼠模型中,我们引入了由此产生的 CAR 转基因 Treg 来测试它们的治疗潜力在体内。这包括评估脱色,以及皮肤中 Treg 和黑素细胞的持久性。抗原特异性 Treg 的过继转移可能会提供一个令人兴奋的机会来停止色素沉着并补充新兴的治疗方法,例如修饰诱导型 HSP70 以耐受树突状细胞 (DC) 和 Janus 激酶 (JAK) 抑制剂以减轻 T 细胞活化 ( 36 , 37 ).
材料和方法
学习规划
本研究的目的是研究 GD3 CAR Tregs在人类白癜风小鼠模型中阻止色素脱失的功效。我们在体外极化和扩增 Tregs ,然后使用 CAR 构建体生成抗原特异性 Tregs,以生成具有高转导效率的 GD3 CAR Tregs。为了确定体外效应,我们在存在未转导的 Treg 和 GD3 CAR Treg 的情况下,将人 HLA-A2 +黑素细胞与酪氨酸酶反应性效应 T 细胞共同培养。对于体内疗效后,我们过继地将未转导的 Tregs、GD3 CAR Tregs 和 HBSS 载体转移到易患白癜风的人源化小鼠模型中,对动物进行脱色分析,同时通过免疫组织学和多重细胞因子分析评估 CAR Tregs 的局部和远距离效应。对来自白癜风患者的人皮肤进行染色以检测损伤、损伤周围和非损伤皮肤上的 GD3 表达。
体内实验的样本量是根据先前研究的统计功效计算和过去对白癜风 h3TA2 小鼠模型的经验确定的。对于体内研究,小鼠被随机分配到性别组,研究人员对实验条件和进一步分析不知情。所有实验样本和动物都包括在分析中,不排除异常值。样本量、重复次数和统计方法显示在结果和图例中。
组织采购、细胞培养和试剂
人类病灶周围皮肤组织是在白癜风患者的知情同意下获得的,该患者在伊利诺伊州梅伍德洛约拉大学医学中心的皮肤科诊所就诊(补充表 1)。研究得到了机构审查委员会的批准,符合赫尔辛基宣言中描述的原则。幼稚小鼠 CD4 + T 细胞和 CD4 + FoxP3 +Tregs 在补充有 10% FBS、1X 非必需氨基酸(Corning)、50 U/ml 青霉素-链霉素(ThermoFisher Scientific)、1 mM 丙酮酸钠(GibcoLife Technologies)、10 mM HEPES(Gibco)的 RPMI 培养基中培养Life Technologies)和 50 µM β-巯基乙醇(Sigma Aldrich)。人类黑色素细胞在人类黑色素细胞生长补充剂 2(Thermo Fisher Scientific)中培养,添加到培养基 254(Thermo Fisher Scientific)中,加入 10 mM L-谷氨酰胺(Thermo Fisher Scientific)和 1X 抗生素抗真菌剂(Thermo Fisher Scientific)。如所述( 38 )产生兔抗GD3 CAR血清和稳定的GD3 CAR病毒产生细胞。将稳定的GD3 CAR病毒产生细胞(VPC)维持在上述T细胞培养基中用于病毒产生。
在体外分离幼稚 CD4+ T 细胞并极化为 CD4+FoxP3+ Tregs
使用 EasySep 小鼠幼稚CD4 + T 细胞分离试剂盒 (StemCell Technologies)从 8-10 周龄 B6.Cg- Foxp3 tm2Tch /J(“FoxP3 eGFP”) 报告小鼠 (Jackson Laboratories)的脾脏中分离幼稚小鼠 CD4 + T 细胞遵循制造商的协议。这些小鼠共同表达增强型绿色荧光蛋白 (eGFP),它仅限于 T 细胞谱系,主要是 CD4 + T细胞群。初始 CD4 + T 细胞极化为 CD4 + FoxP3 +在Dynabeads Mouse T-Activator CD3/CD28 (Thermo Fisher Scientific) 存在下使用 30 ng/ml 人转化生长因子 β (TGF-β) (eBioscience),珠与细胞的比例为 1:1,300 国际单位/ ml(IU/ml) 重组人白细胞介素 2 (rhIL-2) (NIH, Bethesda, MD) 5 天。人 TGF-β 用于极化鼠类 Treg,因为小鼠和人 TGF-β 具有 99% 的序列同源性,具有高跨物种活性 ( 39 , 40 )。人 IL-2 被用作人 IL-2 有效地刺激小鼠 IL-2 受体,而小鼠 IL-2 不会引发与人 IL-2 受体的有效结合 ( 41 , 42 )。
GD3 CAR转导小鼠 Treg 的产生
24 个孔的非组织培养板涂有 10 μg/ml 反连接蛋白(Takara Bio USA Inc.)。如所述( 43 )生成对GD3反应的基于MFG逆转录病毒载体的第二代CAR构建体(sFv-CD28/TCRζ )。将来自 Phoenix E 逆转录病毒生产细胞 ( 43 ) 的条件培养基上清液转移至由编码 GD3 CAR 的病毒 (80% 汇合)( 43 , 44 ) 组成的逆转录酶包被板中,并以 2,000xg 离心。小心去除上清液并激活 CD4 + FoxP3 +将Treg 转移到带有额外病毒上清液、5 μg/ml 硫酸鱼精蛋白 (Sigma Aldrich) 和 300 IU/mL rhIL2 的后连接蛋白包被板上。将板以 1,000xg 离心,然后如上所述与完全 T 细胞培养基和小鼠 T-Activator CD3/CD28 珠和 rhIL-2 一起温育。然后重复转导以提高转导效率。在流动分析之前,用 CD3/CD28 珠、30 ng/ml 人 TGF-β 和 rhIL-2 重新激活转导的 Treg 2 天。
流式细胞术
在表面染色之前,根据制造商的说明,将细胞与小鼠Fc 块 (BioLegend) 和LIVE/DEAD 可固定近红外死细胞染料 (Thermo Fisher Scientific) 一起孵育。直接标记抗体的表面染色包括 BUV395 标记的抗小鼠 CD3 克隆 145-2C11 (BD Biosciences) 和 BV421 标记的抗小鼠 CD4 克隆 GK1.5 (BioLegend)。在Treg 报告小鼠的 FoxP3 启动子下表达的 eGFP 标记以及 BB700 标记的大鼠抗小鼠 CD25 克隆 PC61 (BD Biosciences) 用于鉴定 Treg。通过抗兔别藻蓝蛋白 (APC) (Invitrogen) 抗体检测到的未标记抗 GD3 CAR 兔血清用于评估转基因 Treg 的 CAR 表达。使用 BDFACSymphony 流式细胞仪和 FlowJo v10.3.0 软件(FlowJo LLC,OR,USA)分析染色的细胞。
体外共培养实验
在体外共培养实验之前,通过使用 FITC 标记的 BB7.2 对人 HLA-A2进行免疫荧光染色来鉴定HLA-A2 +黑素细胞。用酪氨酸酶反应性 h3T 效应 T 细胞 (Teffs) ( 29 )和未转导或 GD3 CAR 转导的抑制因子接种人 HLA-A2 +新生儿包皮黑色素细胞(Mf0887,P6)和 HLA-A2- 腹壁增生性皮肤黑色素细胞(Ms18001,P6)Tregs 以10:1:1 的效应器与目标与抑制器的比例持续 36 小时。Teff:Tregs 比率用于模拟 T 细胞亚群的自然发生,因为 Tregs 占总 T 细胞群的5-10% 共培养物一式三份接种并使用 IncuCyte®孵育Caspase-3/7Red Apoptosis Assay Reagent (Sartorius)。使用 IncuCyte 活细胞分析系统 (Sartorius) 每 3 小时拍摄一次图像,一式三份。根据制造商的方案,将上清液保存用于小鼠 IFN-γ(R&D systems,Minnesota,MN)和 IL-10ELISA 测定(Mabtech AB,Stockholm,Sweden)。通过使用 Adobe Photoshop (AdobeSystems, San Jose, CA) 相对于对照孔量化活细胞来检查细胞毒性。
过继性 Treg 转移
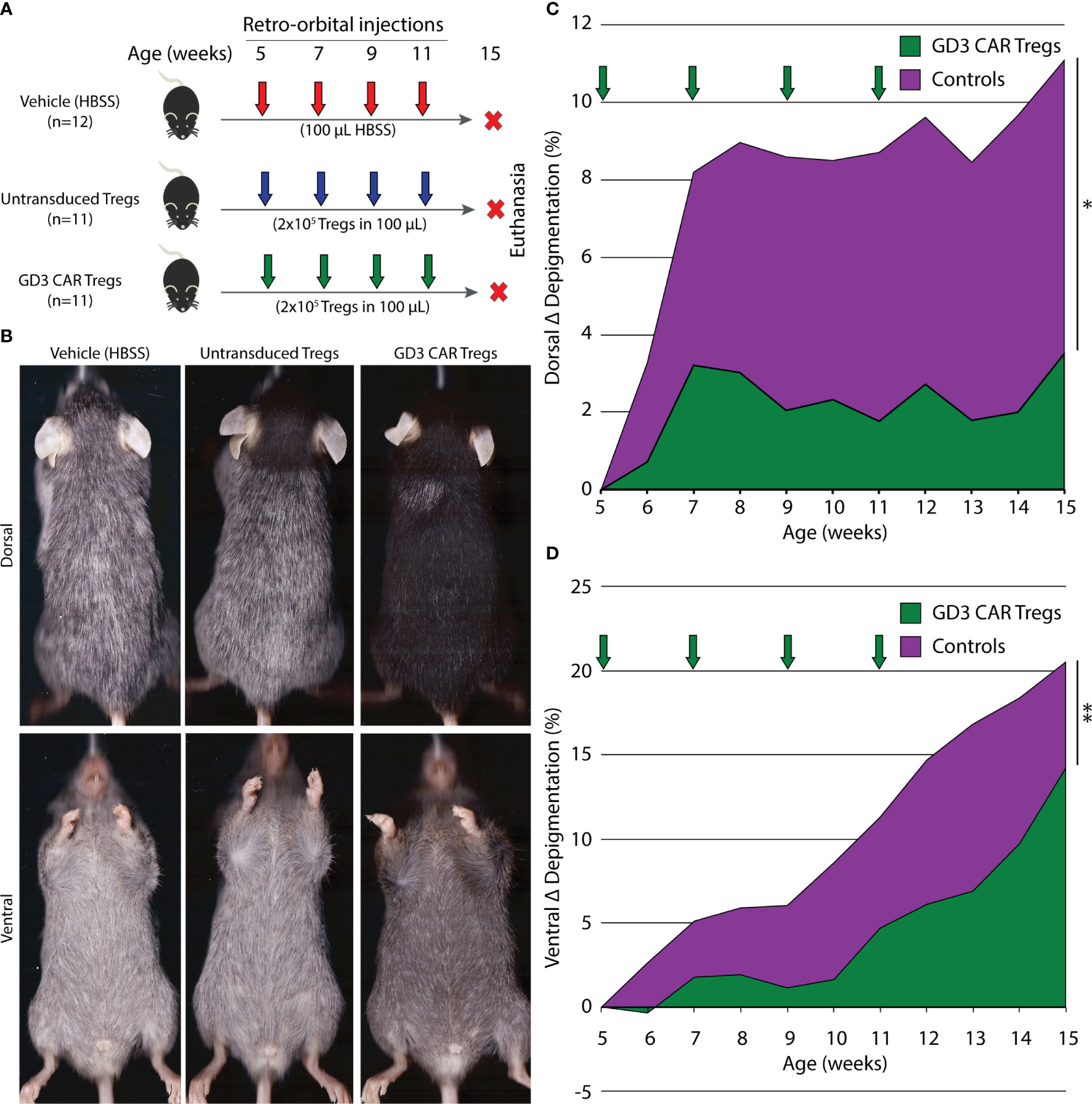
根据西北大学机构动物护理和使用委员会 (IACUC )批准的方案,按照关于护理和使用美国国家研究委员会概述的实验室动物。每只动物(n = 11; 6♂,5♀)或 2x10 5 GD3CAR Tregs/每只动物(n = 11;6♂,5♀)对小鼠进行眶后给药,或用载体治疗( HBSS)单独(n = 12;6♂,6♀)四次,每两周一次,从 5 周龄开始。确定了过继转移的 Tregs 的数量,以便与我们早期的研究 ( 21 ) 进行比较,其中2x105 个多克隆 Treg 控制人酪氨酸酶 TCR 转基因 HLA-A2 (h3TA2) 小鼠模型中 3-9 周龄小鼠的色素脱失。所有组在整个实验过程中每周 3 次接受低剂量的重组人 IL-2(3,000IU),以促进过继转移的 Tregs 的体内刺激(45 , 46)。动物被维持15周并人道地安乐死。实验在 6 个不同的时间点开始,包括来自不同窝的小鼠。幼稚 T 细胞被极化和转导以产生 CAR Treg,每次最多 4 只小鼠/组。随着时间的推移监测每个实验的脱色情况,将结果汇总以进一步证实结果。皮肤活检、脾脏、脑、回肠、淋巴结保持在最佳切割温度 (OCT) 下,并储存血清用于细胞因子分析。
脱色分析
从 5 周到 15 周龄,小鼠在异氟醚麻醉下每周在平板扫描仪(Hewlett-Packard,Plo Alto,CA)上进行扫描。使用 AdobePhotoshop 软件 (Adobe Systems) 测量腹侧和背侧亮度以计算脱色,如前所述 ( 47 )。随时间绘制脱色图,并通过时间调整的曲线下面积 (AUC) 确定统计显着性。使用梯形规则计算代表从治疗开始的脱色变化。没有对缺失数据进行插补,每只小鼠的 AUC 除以可用数据的总周数减 1。Wilcoxon秩和 (WRS) 检验用于比较组间的时间调整 AUC。
免疫组织学
使用 OCT 化合物 (Sakura Finetek) 在干冰上冷冻小鼠和人类皮肤样品。切割了 8 µm 冷冻切片(徕卡)。对于 FoxP3/CD3 染色,使用真核转录因子缓冲液组 (BioLegend) 对切片进行多聚甲醛固定和透化。切片用 SuperBlock (ScyTek Laboratories, Logan, UT) 处理。小鼠 CD3(Biolegend) 的 PE 标记抗体145-2C11 和小鼠 FoxP3 (BioLegend) 的Alexa Fluor 488 标记抗体 MF-14 用于双重染色程序,然后是 4',6-diamidino-2-phenylindole (DAPI ) (BD Biosciences) 核染色。对于其他组织染色,将小鼠和人类皮肤切片固定在冷丙酮中。小鼠皮肤切片用 SuperBlock 封闭,然后与针对 TRP-1 的抗体 H-90(Santa Cruz Biotechnology,Dallas, TX),然后是 Alexa Fluor 555 标记的驴抗兔抗体 (abcam),或 PE 标记的MB3.6 到 GD3 (Santa Cruz Biotechnology),或 PE 标记的抗体 YGITR 765 到糖皮质激素诱导的 TNF 受体 (GITR) (Biolegend) ),或 AF488 标记的抗体 B56 至Ki67 (BD Biosciences),均随后进行 DAPI 核染色。用 10% 正常人血清(Gemini Bio Products,West Sacramento,CA)封闭人皮肤切片,然后用 Ta99 孵育 TRP-1(BioLegend)或 R24 孵育 GD3(Abcam,Cambridge,UK)。两者均由HRP 缀合的山羊抗小鼠 IgG 抗体(AgilentDako,Santa Clara,CA)检测。这些染色是使用 AEC 底物 (Abcam) 显影的,随后通过在 Mayer 苏木精 (Sigma-Aldrich) 中孵育并在 Scott 自来水 (Sigma-Aldrich) 中发蓝来检测细胞核。载玻片在 Revolve 显微镜(Echo Laboratories)上成像。使用 AdobePhotoshop 软件对细胞进行量化。
细胞因子分析
细胞因子分析包括来自体外抑制测定(IncuCyte 实验)、共培养 36 小时后收集的上清液,以及来自 HBSS 载体(n = 11)、未转导(n = 10)和 GD3 CAR Tregs(n = 9) 处理的小鼠组。小鼠干扰素 γ (IFN-γ)、肿瘤坏死因子 (TNF-α)、IL-4 和IL-10 的检测通过使用针对这些小鼠细胞因子的定制 V-Plex 面板 (Meso Scale Diagnostics, LLC) 进行根据制造商的说明。在MESO Quickplex SQ120(Meso Scale Diagnostics,LLC)上获取数据,并使用 Prism 版本 8.3.0(GraphPad 软件)进行分析。
统计分析
使用 GraphPad Prism 8.0 软件 (GraphPad) 和 R 软件进行统计分析。数据显示为具有平均值±标准偏差的条形图和点图。数据通过单因素方差分析 (ANOVA) 方差分析评估治疗组之间的不同方差,并进行事后 Tukey-Kramer 比较。确定体外免疫抑制的统计学意义, 双向 ANOVA 与对齐秩变换一起使用,然后使用 Tukey 方法进行多对比较测试。对于脱色,使用梯形规则计算时间调整的 AUC,代表治疗开始后脱色的变化。没有对缺失数据进行插补,每只小鼠的 AUC 除以可用数据的总周数减 1。WRS 检验用于比较组间的时间调整AUC。统计显着性表示为 *p < 0.05、** p< 0.01、*** p < 0.001 或**** p < 0.0001。
结果
GD3 由包括黑素细胞在内的病灶周围上皮细胞表达
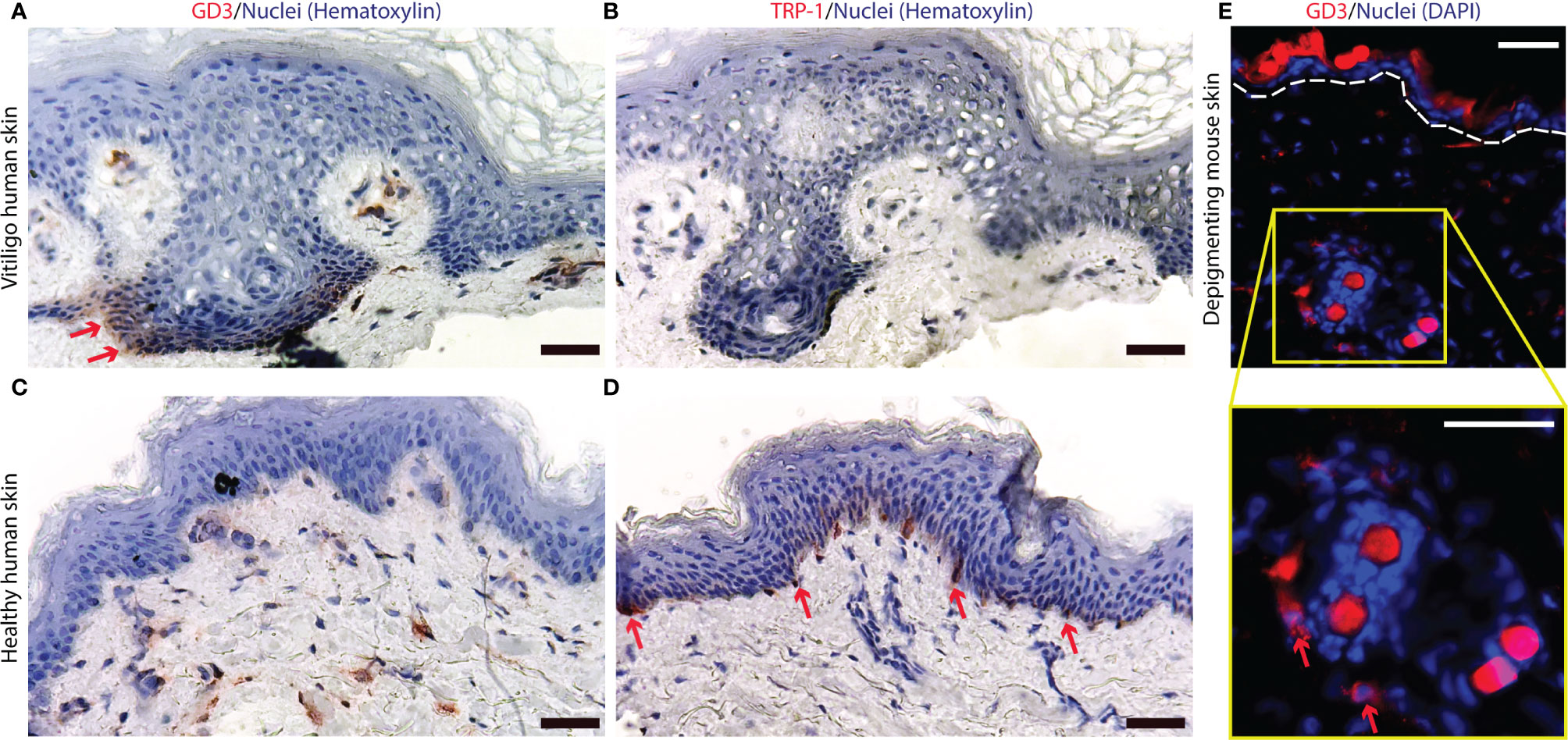
据报道,过表达 O-乙酰化 GD3 可有效去除白斑皮肤的色素 ( 31 , 38 )。这促使我们评估GD3 在皮肤活检中的表达本身,这些活检取自积极脱色皮肤的病灶周围活检。在人白癜风病灶周围表皮中观察到 GD3的显着表达(图 1A),而显示的边界活检切片中没有黑素细胞(图 1B)。图 1A和1B中的空泡化经常在白癜风皮肤中观察到,并且已被其他人提出作为白癜风的指征 ( 48 , 49 )。在健康对照皮肤中未观察到表皮 GD3 表达(图 1C )而黑色素细胞在该组织中很容易检测到 (图 1D )。类似地,在脱色 h3TA2 小鼠皮肤中发现了 GD3 表达(图 1E)。
图1
图 1神经节苷脂 D3 在人和小鼠来源的脱色皮肤中表达。(A) GD3 的表达和(B)缺乏黑色素细胞,表现为缺乏 TRP-1+ 染色,在来自病灶周围皮肤的人类脱色素表皮中发现。而(C)有限的 GD3 表达和(D)丰富的 TRP-1 表达显示在健康对照皮肤中。(E)在脱色小鼠皮肤中,在毛囊周围和表皮附近观察到表达 GD3 的细胞。GD3 表达细胞的代表性染色显示为红色,细胞核为蓝色,在自发荧光小鼠毛发周围(比例尺 = 50μm)。
使用GD3 编码的 CAR 构建体实现了 Treg 的高病毒转导
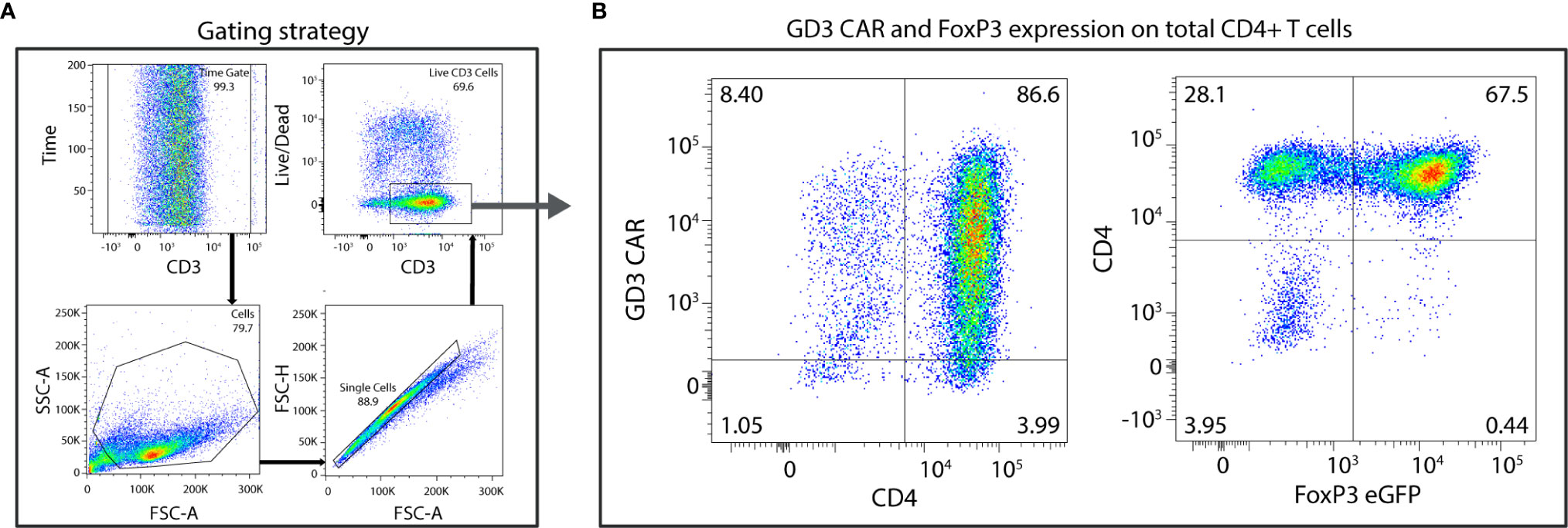
为了生成在需要时参与抑制活动的治疗性 Treg,我们生成了 FoxP3 + CD4 + T细胞并转导它们以表达 GD3 反应性 CAR。在一个有代表性的例子中,从 3x10 8个脾细胞中分离出大约 1.5x10 6 个幼稚CD4 + T 细胞,在存在 TGF-β 的情况下保持,并成功地极化和放大到每只供体小鼠大约1.6x10 7 个Tregs。逆转录病毒转导TGF-β 极化的初始 CD4 + T细胞,并通过流式细胞术评估 GD3 CAR 表达。门控策略如图 2A所示,其中 86.6% 的总 CD4 +用GD3 CAR 构建体成功转导了 T 细胞(图 2B)。进一步扩增后,64 ± 3.5% 的转导细胞是 FoxP3 + Tregs。从4x10 6 FoxP3 + Tregs、2.1x10 7 GD3CAR-expressing、FoxP3 + Tregs的初始预扩展和转导池中产生。大多数产生的 CAR 转导的 Treg 预计将作为免疫抑制 T 细胞发挥作用,并发挥局部免疫抑制功能。我们接下来在体外测量了 GD3CAR Treg 功能。
图 2
图 2观察到表达 GD3 CAR 的 Tregs 的高转导效率。CD4 + FoxP3 +细胞,从幼稚的 CD4 + T 细胞极化,使用 GD3 CAR 编码构建体进行转导。(A)门控策略包括一个时间门,然后依次对淋巴细胞、单细胞和活细胞进行门控。(B) 86% 的总 CD4 + T 细胞成功转导以表达 GD3 CAR 构建体,67% 的细胞群表达 FoxP3 +。
抗原特异性增加免疫抑制细胞因子的产生
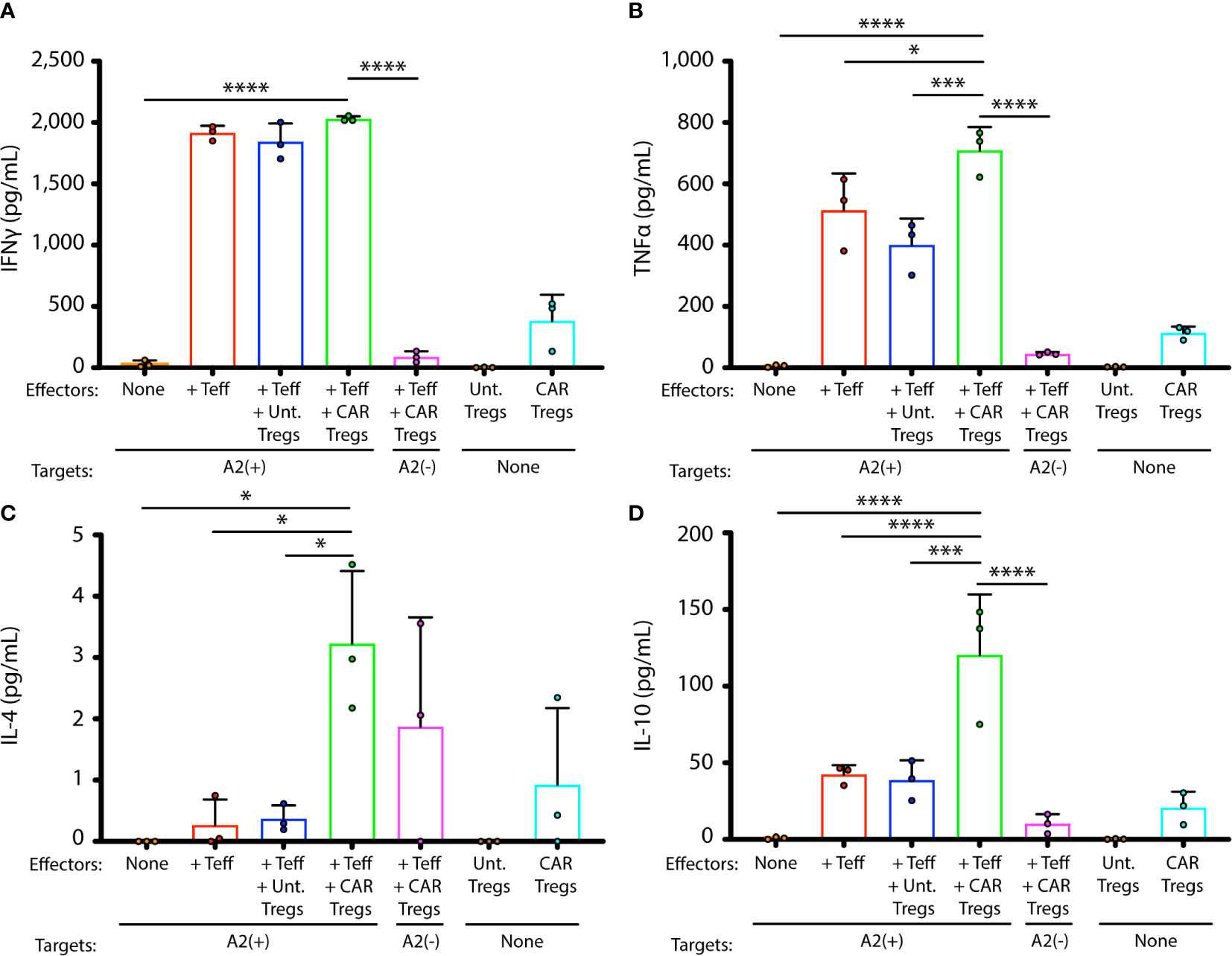
在 GD3 CAR Tregs 或未转导的 Tregs 与酪氨酸酶反应性 Teff 及其 HLA- 的共培养物中测量了与免疫激活或免疫抑制相关的代表性细胞因子 IFN-γ、TNF-α、IL-4 和 IL-10 的产生。匹配的靶标 (1:10:1),在细胞在 IL-2 存在下培养 42 小时后测量浓度(图 3)。这些 Teff 可以识别人类黑色素细胞 ( 29 )。在包含或不包含Treg 的组合中未发现 IFN-γ 产生的显着差异,这表明后者在此 Treg 与 Teff 的比率下对这种细胞因子的产生几乎没有影响(图 3A)。显着更多的 TNF-α(图 3B,p = 0.0005)、IL-4(图 3C, p= 0.03), 并且 IL-10 (图 3D ,p = 0.0005) 是与 CAR Tregs 组合产生的,尽管 IL-4 的总产量一直很低。重要的是,仅在细胞毒性 T 细胞和 HLA 匹配的人类黑素细胞存在时观察到增加的 IL-10 调节细胞因子产生。总之,细胞因子环境表明存在抗原特异性 Treg 时具有更强的免疫抑制能力,由活化的效应 T 细胞刺激。为了测量这种细胞因子环境是否可能转化为对黑素细胞靶细胞在体外细胞死亡的更大保护,我们接下来测量了黑素细胞、Teff 和 Tregs 共培养物中持续的靶细胞活力。
图 3
图 3 GD3CAR Tregs 在活化的 T 细胞存在下产生免疫抑制细胞因子。在存在和不存在未转导或 CAR 转导的 Treg 的情况下,在来自黑素细胞靶标和 HLA-A2 限制性 Teff 的共培养物的上清液中测量细胞因子。每个共培养的细胞因子浓度,在三次共培养中测量,显示为(A) IFN-γ、(B) TNF-α、(C) IL-4 和(D) IL-10。通过单向方差分析测试进行统计分析,然后进行 Tukey's post-hoc 测试以进行多重比较。*p <0.05; **p < 0.01; ***p < 0.001;****p< 0.0001。
抗原特异性增加体外Treg 的免疫抑制活性
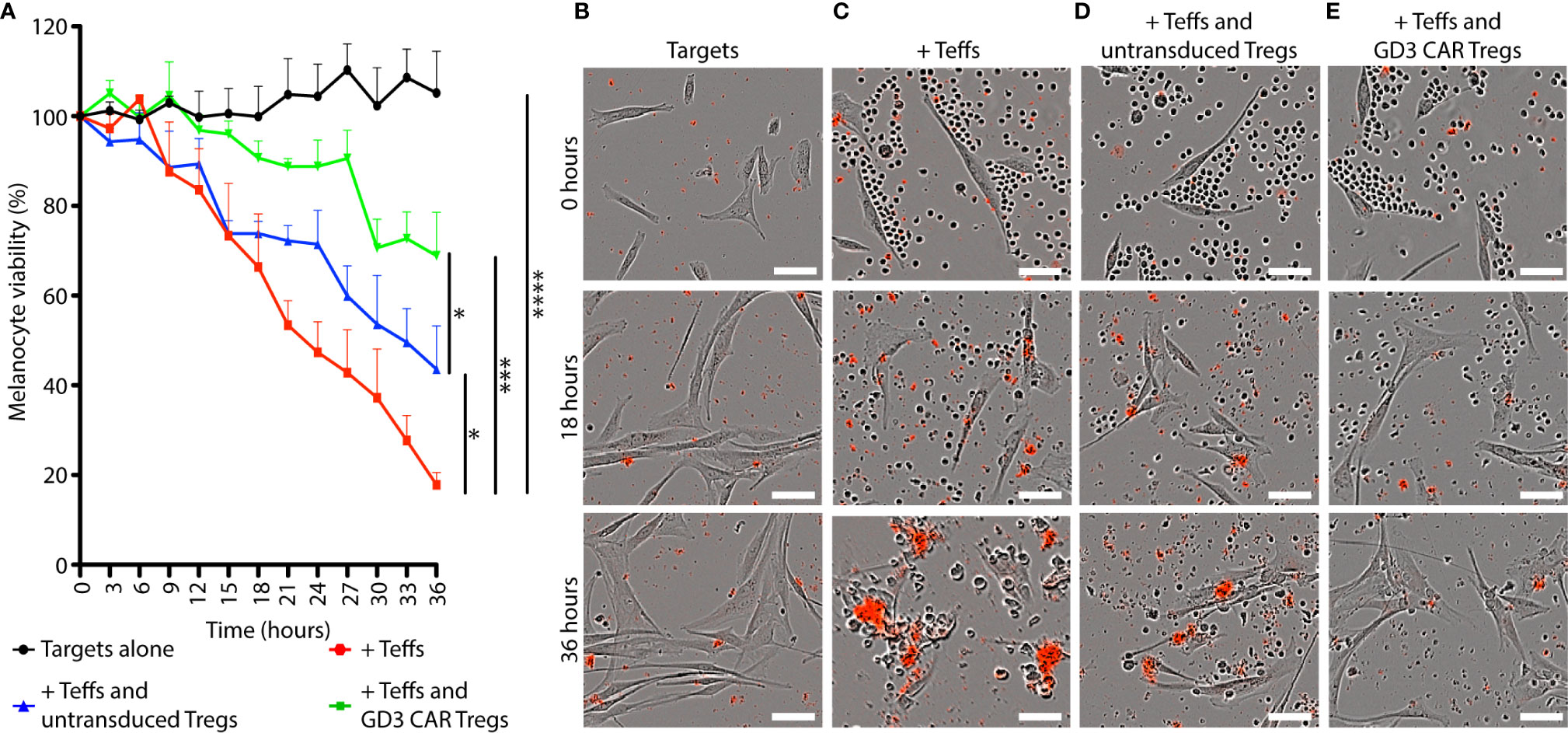
Tregs通过细胞因子、细胞间接触或旁观者效应抑制常规 T 细胞( 50 )。为了测量由此产生的抑制活性,我们在体外评估了靶标、Teffs 和 Tregs 共培养36 小时的持续黑素细胞活力。图 4A显示了靶向 HLA-A2 +的活力目标不同组合中的人类黑素细胞,Teffs 和 Tregs 1:10:1。在没有 Teff 细胞的情况下,存活目标的数量随着时间的推移略有增加。相比之下,在存在效应 T细胞 36 小时后观察到 82.2% 的细胞毒性 (p < 0.0001)。随着时间的推移,未转导的 Tregs 提供了 35.8% (p = 0.02) 的细胞毒性保护。使用 R 软件进行双向 ANOVA 和对齐秩转换,并根据 Tukey 进行成对事后多重比较测试,以确定在存在 CAR Treg 的情况下,防止了 62.0% 的对黑素细胞的细胞毒性(p = 0.0004)。代表每个细胞组合的图像,包括单独的目标(图 4B)、目标和 Teff(图 4C),以及存在未转导 Treg 的后一种组合(图 4D )或 CAR Tregs (图 4E )在不同时间点同样显示了在包括 GD3 CAR Tregs 的组合中对细胞毒性的大部分抑制。因此,未转导的 Tregs 和 GD3 CAR Tregs 都提供了对黑素细胞活力的显着保护。重要的是,与未转导的 Tregs 相比,GD3 CAR Tregs 提供的保护作用要大得多( p = 0.04 ),这表明抗原特异性对增强免疫抑制有额外的好处。因此,我们接下来探索了 CAR Tregs在体内的治疗效果。
图 4
图 4 GD3CAR Tregs 为黑素细胞提供卓越的体外T 细胞介导的细胞毒性保护。在体外 比较了GD3 CAR Tregs和未转导Tregs的免疫抑制能力。(一) HLA-A2 +人黑素细胞(靶标)在鼠 Teffs 和 Tregs(1:10:1)存在或不存在的情况下的活力随时间变化。(B) HLA-A2 +人类黑色素细胞(C)与鼠 Teffs 结合的代表性图像,以及(D)鼠未转导 Tregs 或(E)GD3 CAR Tregs。死细胞由半胱天冬酶活性形成的红色沉淀物标记。统计显着性由双向方差分析确定,对齐秩变换,然后是 Tukey 的成对多重比较检验。*p < 0.05; **p <0.01; ***p < 0.001;****p < 0.0001(比例尺 = 50μm)。
抗原特异性 Treg 增强 h3TA2 小鼠的免疫抑制
为了评估 CAR Treg 在进行性脱色模型中的抑制活性,我们测量了从 5 周龄开始的自发脱色 h3TA2 小鼠的脱色。脱色在出生后不久开始,动物在 23 周内显示一半最大脱色 ( 51 )。小鼠接受过继转移的未转导 Tregs、转导 GD3 CAR Tregs 或载体,每两周一次,持续 11 周,如图 5A中所述。图 5B显示了注入未转导 Treg、GD3 CAR Treg 或车辆的动物的代表性背侧和腹侧图像. Wilcoxon秩和(WRS)检验用于比较组间的时间调整曲线下面积(AUC)。载体和未转导的 Treg 对照组的结果没有差异(背侧 p = 0.97,腹侧 p = 0.88)。因此,将载体和未转导的 Treg 组合并,并与 GD3 CAR Treg 治疗组进行比较。在 WRS 测试的单侧 t 近似值中,CARTreg 治疗的小鼠 (n = 11) 在 15 周的观察期内,背部色素沉着的 AUC 下降了 73.0% (p = 0.028)。腹侧色素脱失发生得更快,并单独评估。在这里,CAR Treg 治疗组 (n = 11) 在随访期间 (图 5C )的脱色显着延迟,导致 CAR Treg 治疗小鼠的 AUC(p = 0.006) 降低了 60.5% (p = 0.006)。图 5D)。每只小鼠的单个背侧和腹侧脱色值显示在补充数据文件中(补充图 1)。CAR Tregs 增强的疾病控制可能是由于 GD3 表达对抑制活性的局部激活和现场活化 Teff 的存在。为了评估这一点,在仅用载体 (n = 11)、未转导的 Tregs (n = 10) 处理的小鼠的血清样本中测量了 IFN-γ、TNF-α、IL-4 和 IL-10 的血清细胞因子含量的变化,或 GD3 CAR Tregs (n = 9)。结果的细胞因子水平在终点的各组之间非常一致(补充图2)。该结果支持Tregs(包括CAR Tregs)可能在免疫活性区域被优先激活的概念。整个实验过程中未观察到不良事件,任何组的小鼠在安乐死的内脏检查中均未发现异常。我们接下来探讨了减少的脱色是否伴随着黑素细胞的持续存在和 T 细胞群的变化。
图 5
图 5 GD3CAR Tregs 为易患白癜风的小鼠提供显着的色素脱失保护。(A)实验大纲在 6 个不同时间点开始,显示易患白癜风、h3TA2、仅用载体治疗的小鼠 (n = 12),或通过过继转移未转导的 Tregs (n = 11) 或 GD3 CAR Tregs (n = 11)。过继转移从 5 周龄开始,每两周持续一次,直到 11 周龄。从 5-15 周龄开始每周测量一次脱色情况。(B)来自 HBSS 载体、未转导 Treg 和 GD3 CAR Treg 治疗组的小鼠在 15 周龄时的代表性背侧和腹侧扫描。(C)背部和(D)量化的脱色整个实验的腹侧。Wilcoxon 秩和 (WRS) 检验用于比较组间的时间调整 AUC。箭头:治疗时间。* p < 0.05 ; ** p < 0.01。
黑素细胞在 GD3 反应性 CAR Tregs 的存在下受到保护
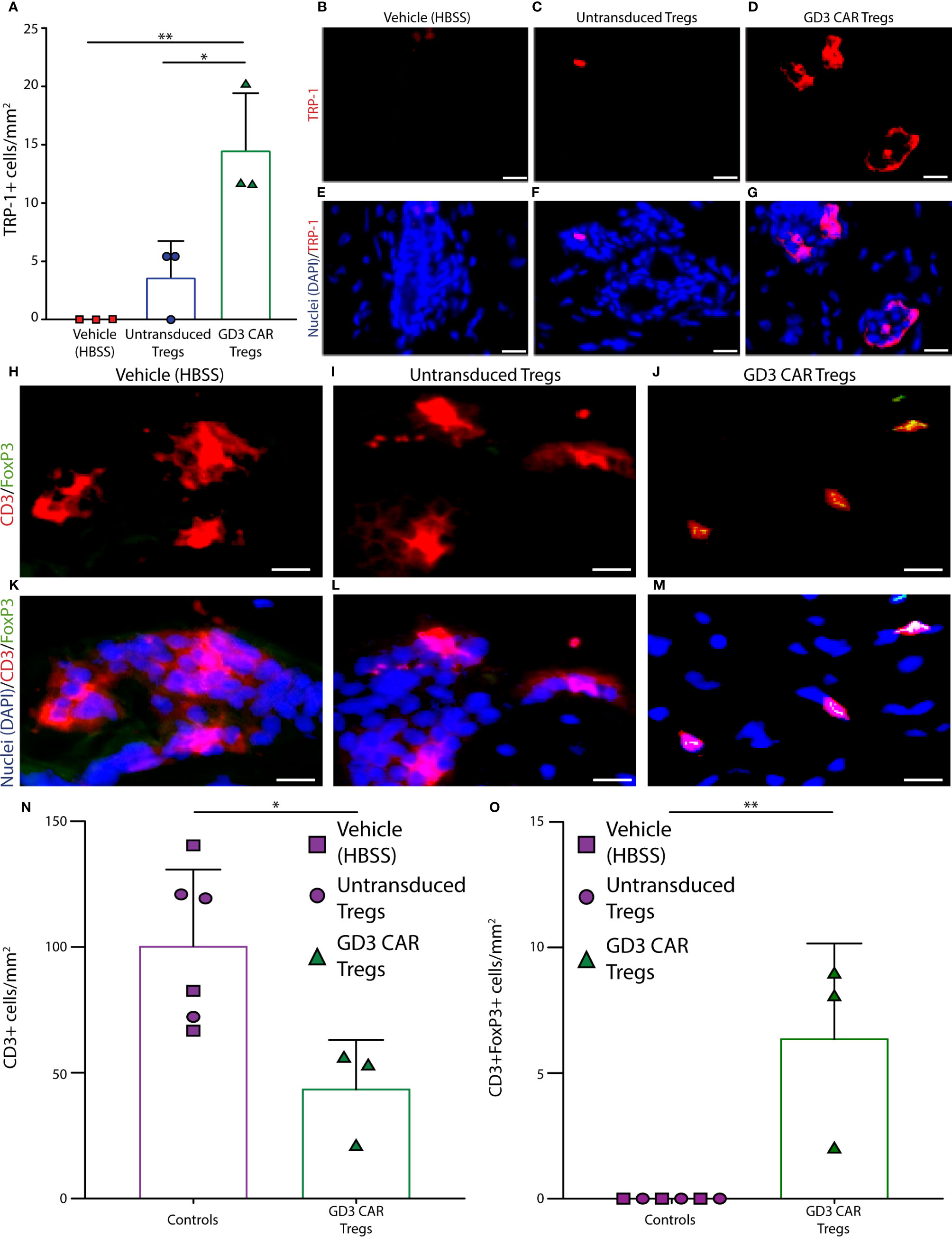
使用针对 TRP-1 的抗体评估小鼠背部皮肤活检的黑素细胞丰度,如图 6所示。如图 6A所示对黑素细胞进行量化,其中来自载体处理的小鼠(每组 n = 3)的皮肤样本显示黑素细胞完全丧失。来自未转导的 Treg 治疗小鼠(每组 n = 3)的皮肤仅显示少数剩余的黑素细胞,并且进行单向方差分析,然后进行 Tukey的事后测试,以证明来自 CAR Treg 治疗小鼠的皮肤含有显着更多的数量与用未转导的 Treg 处理的小鼠(p = 0.025)和载体处理的对照(p = 0.006)相比,黑色素细胞的数量。载体处理的、未转导的 Treg 处理的和 CAR Treg 处理的小鼠的 TRP-1 染色的代表性图像显示在图 6B-D ,并在图 6E-G中分别覆盖了 DAPI 核染色。在检查 GD3 表达时发现了类似的结果。GD3 表达细胞的定量显示,与载体 HBSS 处理的小鼠(p = 0.003)或输注未转导的 Treg 的小鼠(p = 0.003)相比,输注 CAR-Tregs 的小鼠维持显着更多的 GD3 表达细胞(补充图 3)。该观察结果支持表达GD3的细胞没有经历在载体处理或未转导的Treg处理的小鼠中观察到的细胞毒性的概念。这种确认性黑色素细胞定量主要对应于图 6中所示的体内数据,证明了 CAR Tregs 改进的抑制能力。为了解释色素沉着和黑素细胞维持的差异,我们接下来将这些数据与每个治疗组中的 Treg 浸润进行比较。
图 6
图 6在GD3 反应性 CAR Tregs 的存在下,黑色素细胞免受h3T 细胞毒性 T 细胞的侵害。使用针对 TRP-1 的抗体评估小鼠皮肤的黑素细胞存在。(A)黑素细胞的量化,以及来自(B) HBSS 载体、(C)未转导的 Tregs 和(D) GD3CAR Tregs 处理的小鼠(每组 n = 3)的TRP-1 染色的代表性图像,与( E-G)显示了相应的叠加层,包括蓝色的 DAPI 核染色。还使用针对 CD3ε 和 FoxP3 的抗体以及来自(H)载体对照、(I)未转导的 Treg 和(J)显示了 GD3 CAR Tregs 给药的小鼠。代表性样本用于量化 CD3 + T细胞(红色)、FoxP3 +细胞(绿色)和双阳性 Treg;(K-M)还显示了带有 DAPI(蓝色)的相应叠加层。分别显示了(N) T 细胞和(O) Tregs的皮肤染色量化 ± SD (每组n = 3)。通过非参数t检验进行统计分析。* p < 0.05,** p < 0.001(比例尺 = 20 μm)。
CARTregs 倾向于皮肤中的 GD3 表达细胞
为了了解 Treg 活性是否与现场免疫抑制性 T 细胞的丰度相关,使用 CD3 和 FoxP3 抗体评估小鼠皮肤组织的 T 细胞浸润。图 6还显示了来自载体对照组的皮肤示例,以及来自用未转导或 CAR Treg 治疗小鼠治疗的小鼠的样品。在图 6H-J中,相同组的Tregs 被确定为 CD3ε + FoxP3 +细胞,分别在图 6K-M中覆盖有 DAPI 核染色。CD3 +细胞和 CD3 + /FoxP3 +Treg丰度被量化为每个治疗组的平均值±SD(每组n = 3)。在 Tukey事后检验后的单向 ANOVA 中,与 CAR Treg 治疗组相比,对照组在终点浸润 CD3ε + T 细胞的平均数量是 2.3 倍(p = 0.02)(图6N )。在任一对照组中均未检测到(剩余)CD3 + FoxP3 + Treg,而在过继转移 10 周后,CAR Treg 治疗小鼠的皮肤组织中仍可检测到一些 CD3 + FoxP3 + Treg(图 6O)。通过 GITR 表达评估 Treg 数量,与用未转导 Treg (p = 0.0059) 或单独载体 (p = 0.0089) 治疗的小鼠相比,在 CAR Treg 治疗小鼠的皮肤中再次观察到 Treg 数量增加,但仍有组间增殖的 GITR + Ki67 +细胞的丰度没有差异。这表明 Treg 丰度的差异可能由 CAR Treg 治疗小鼠皮肤中 Treg 的流入增加或流出减少来定义(补充图 4)。尽管如此,CAR Treg 治疗小鼠中 Treg 丰度的增加可能解释了 CAR Treg 抑制活性的提高,并表明局部抗原识别支持维持 Treg 的存在。图 7)。总之,数据显示抗原特异性延长了过继转移的 Treg 的抑制活性。
图 7
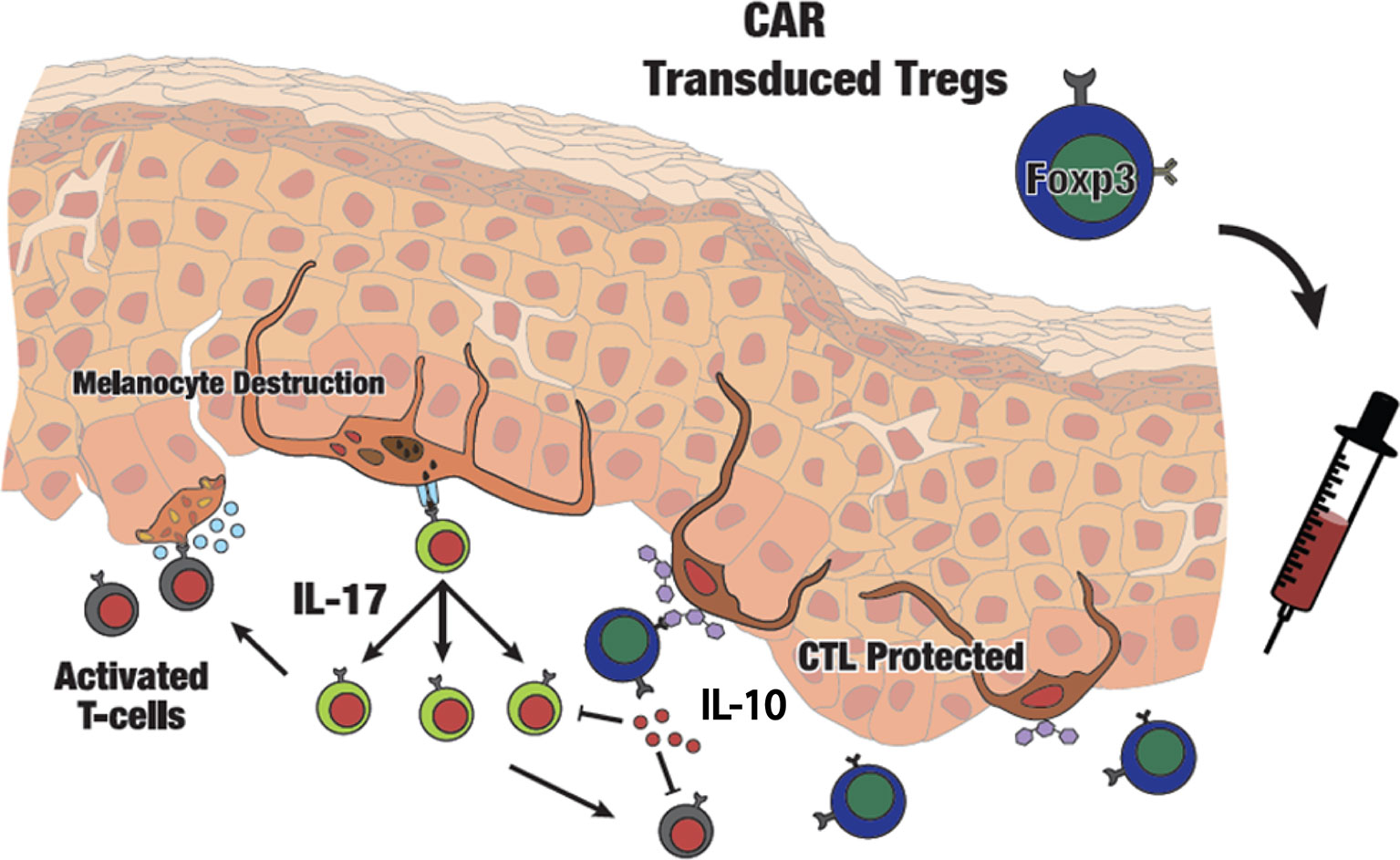
图 7白癜风中 CAR Tregs 过继转移的示意图。自身免疫性黑素细胞破坏由细胞毒性 T 细胞介导,细胞毒性 T 细胞通过应激黑素细胞分泌的自身抗原激活。升高的 IL-17 会促进皮肤中的炎症环境。注入的 GD3 特异性 CAR Treg 将渗入皮肤并对自身免疫活性部位表达的 GD3 作出反应,抑制现场细胞毒性 T 细胞,并在白癜风病灶周围皮肤提供局部免疫耐受。
讨论
在这里,我们描述了工程化 GD3 CARTregs 为自身免疫性白癜风提供抗原特异性免疫耐受的治疗潜力。与传统的免疫抑制药物、生物制剂、烷化剂和抗代谢物相比,Treg 可以提供更高的特异性和复杂的治疗益处,并恢复各种自身免疫性疾病的免疫耐受性( 20 )。当使用多克隆 CD4 + CD25 + FoxP3 + Tregs在各种小鼠模型(包括白癜风)中加强免疫耐受时,临床前研究提供了有希望的结果( 21 )。建立了一个强大的人类 Treg 分离方案,以对纯度 >90% 的 Treg 进行分类,以及离体扩展协议的开发是为了从单个供体获得超过3x10 9 个Treg,类似于此处使用的协议 ( 52 )。总体而言,这项和其他 I 期临床研究为围绕 Treg 免疫疗法的隔离和扩展提供了答案 ( 53 – 56 )。然而,迄今为止,多克隆 Treg 转移的功效并不是不言而喻的。
在白癜风中,一些研究报告循环 Treg 数量没有显着差异 ( 18 , 57 ),而其他研究报告白癜风患者和健康血液之间的 Treg 丰度存在差异 ( 15 , 58 )。尽管循环 Treg数量可能会有所不同,但在患者皮肤中 Treg 局部缺乏的报告中发现了更多的一致性,这可能是白癜风中不受控制的外周免疫反应和进行性色素脱失的原因 ( 12 , 18 , 57 )。
与多克隆 Treg 相比,胰岛特异性 Treg 的过继转移在阻断 1 型糖尿病进展方面优于多克隆 Treg ( 20 , 59 , 60 )。不幸的是,当胰腺破坏接近完成时,通常会在患者中检测到糖尿病,而其他情况可能更适合在临床环境中接受基于 Treg 的治疗。临床前研究同样表明抗原特异性 Tregs 在移植过程中的卓越功效 ( 61 – 64 )。
在白癜风的h3TA2小鼠模型中,新的Teff不断攻击黑素细胞,从而反映了持续活跃的疾病。在这些小鼠中,Tregs 可能在治疗期间有效,而当过继转移停止时,色素脱失会恢复。实际上,预计过继性 Treg 治疗在活动性疾病期间最有效。然而,在人类患者中,疾病进展期穿插着不活动期,这为黑素细胞干细胞提供了分化和重新填充脱色病变的机会。使用 JAK 抑制剂治疗以抑制 Teff 活性的患者可能会出现色素沉着,但可能需要通过紫外线进行支持性治疗(65)。对于白癜风的间歇性治疗,储存自体 GD3 CAR Tregs 以备后用将是有益的 ( 66 )。
凭借其抗原特异性,这些 Treg 可能提供更好的安全性并降低全身免疫抑制的风险。这得到了增加的 IL-10 分泌的支持,在 Tregs 在存在活化 Teff 的情况下发挥作用,如图所示。在比较表达第一代和第二代 CAR 的 Treg 时,表达具有 28ζ 共刺激结构域的第二代 CAR 的细胞的 IL-10 含量最多(67)。用于转导我们论文中包含的Treg 的 CAR Treg 构建体在通过 CAR 激活后同样显示出显着的 IL-10 产生,这可能解释了这些 GD3 CAR Treg在体外与未转导的 Treg 相比具有改善的抑制活性。事实上,TCR 和 CAR 转导也可能产生更有效和更稳定的 Tregs在体内,白癜风的临床应用,这种情况对于研究抗原特异性 Tregs 的优越性具有重要的优势,因为它提供了几种与疾病相关的靶抗原 ( 68 )。实验小鼠的培养物和血清中 TNF-α 均显着升高。然而,TNF-α 的基因消融与白癜风的发展无关,与野生型 h3TA2 小鼠相比,在脱色方面没有发现差异 ( 21 )。在白癜风中,TNF-α在细胞毒性 T 细胞 (CTL) 的发育过程中发挥作用,并增强 IFN-γ 的表达,这与模拟白癜风的发育有关 ( 21 , 69 , 70)。最近的研究报告称,TNF-α可能通过激活和诱导体内Treg 增殖来促进抗炎状况( 71 – 73 )。因此,TNF-α 在白癜风中可能具有破坏性和保护性,分别通过促进 CTLs 和刺激 Tregs。虽然TNF-α 耗竭阻止了疾病进展并促进了白癜风的色素沉着,但当 TNF-α 抑制剂用于其他自身免疫性疾病时,5,928 名患者中有 18 名新发白癜风。这为白癜风留下了抗 TNF-α 治疗选择,直到完全避免副作用 ( 74 )。
因此,我们询问抗原特异性 T regs 是否可能为自身免疫性疾病白癜风的治疗提供额外的益处。这种情况有一个复杂的病因 ( 2 ),黑色素细胞丢失是一个共同点,免疫机制普遍对疾病的传播负责。基于 TCR 和 CAR 的 Tregs 具有不同的机制和功能特性。低抗原表达水平足以使基于 TCR 的 Tregs 被激活,而基于CAR 的 Tregs 需要高密度的抗原 ( 75 , 76) 表明可以根据靶组织的抗原表达来选择 TCR 或 CAR。我们进行了重复的Treg 注射,因为我们模型中的小鼠表现出慢性活动性疾病,我们还不知道转基因 Treg 在现场保持活跃多长时间。收养转移的 Treg 的长期命运尚未确定,以更好地了解重复申请的必要性。重要的是,CAR 不受 HLA 限制,并且对其靶标部分具有更高的亲和力 ( 77 )。因此,CAR 构建体可以为患有进行性疾病的患者找到普遍的应用。
我们确定了白癜风中抗原特异性 Treg 的潜在靶标。我们的 GD3 CAR Tregs 在白癜风小鼠模型中保护黑素细胞免受 T 细胞介导的破坏,表达人类 TCR 和匹配的人类 MHC,并能够对人类靶细胞作出反应。重要的是,选择的抗原不需要由在反应中幸免的靶细胞本身表达(78)。
尽管在我们目前的研究中在内脏检查期间没有明显的副作用,但仍然存在一些安全问题,包括如果转导的细胞最终产生效应谱,则可能发生细胞因子释放综合征 (CRS) 或神经毒性 ( 79 )。然而,CRS 在靶向液体肿瘤时比在实体组织中更容易发生 ( 80 )。为了对抗任何潜在的副作用并提高安全性,可以掺入包含基于半胱天冬酶的自杀基因的构建体,以便在必要时使 GD3 CAR Tregs 失活 ( 81 , 82 )。
静脉注射抗原特异性 Treg 的一个限制可能是这些急需的免疫抑制细胞在所需部位显示缺乏 ( 60 )。如果全身应用的Treg 没有按要求响应,可能需要局部注射,或者可以在需要 Treg 吸引全身应用的 Treg 的地方引入 CCR4 Treg 归巢受体配体 CCL22 ( 17 )。当可以识别相关抗原时,这使得皮肤自身免疫性疾病特别适合通过抗原特异性 Tregs 进行过继治疗。事实上,GD3 可能支持角质形成细胞增殖,而 O-乙酰化 GD3 在银屑病皮肤中过度表达 ( 83 , 84)。这些发现表明 GD3 CARTregs 也可能在后一种情况下缓和病变。
迄今为止,目前仅在临床试验中测试同种异体抗原反应性 Treg 以防止器官移植后的排斥反应 ( 60 )。在这里,我们制备并扩展了 GD3 反应性 CAR Tregs,其数量适合于小鼠的过继细胞转移治疗。同样可以在患者身上进行,将治疗性 Treg 冷冻保存以备将来使用 ( 85 – 88 )。过继转移的挑战之一是该技术的成本和可扩展性。这促使了开发现成的“通用 CAR Tregs”的概念,该概念易于适应所有患者。多功能性由结合通用 CAR 和目标细胞的模块提供(89)。总之,此处提供的数据支持使用抗原特异性 CAR Tregs 作为白癜风的过继细胞疗法,以控制脱色和支持白癜风的免疫耐受。
迄今为止,患者对 Treg 输注的耐受性良好。目前正在进行的临床试验的结果可以带来关于最佳 Treg 剂量、预期疗效和任何可能引起的副作用的重要见解,并且可以进一步调整治疗策略以支持持续维持和记忆形成 ( 90 ),改善归巢(91 ),以及根据需要消除过继转移细胞( 92 )的安全措施。可以增强Treg 治疗对白癜风以外的病症的功效和特异性,其中可以鉴定抗原以作为工程化的抗原特异性 Treg 的靶标。体内跟踪将使研究小组能够更好地了解抗原特异性 Treg 的维持和记忆形成潜力。总之,基于 Treg 的疗法具有作为白癜风和其他自身免疫性皮肤病的未来疗法的潜力。
数据可用性声明
研究中提出的原始贡献包含在文章/补充材料中。进一步的询问可以直接联系相应的作者。
道德声明
动物研究由西北大学的机构动物护理和使用委员会审查和批准。已从个人获得书面知情同意书,以发布本文中包含的任何潜在可识别图像或数据。
作者贡献
概念化:CLP 和 ZM。方法论:ZM、CLP、CC 和 DJ。形式分析,ZM。调查、ZM、ERD、JME 和 KP-M。统计分析,AI 和 KC。资源、RPJ、SM 和 AO。写作——原稿,ZM。写作——评论和编辑、ZM、CLP、ERD、YOO 和 JRL。可视化、ZM、ERD、JME 和 SWH。监督,中电。项目管理,中电。所有作者都对文章做出了贡献并批准了提交的版本。
资金
这项研究得到了 NIH RO1s AR057643、CA191317 和 LAM 基金会通过授予 CLP 的既定研究者奖的部分支持。ZM的外国实习得到了哈萨克斯坦共和国教育和科学部的博士学位。Al-Farabi 哈萨克斯坦国立大学的课程。
利益冲突
美国和 PCT 国际专利(Utility&PCT,序列号17/072,939&PCT/US2020/056104)于 2020 年 10 月 16 日以“治疗白癜风的材料和方法”提交。
作者声明,该研究是在没有任何可能被解释为潜在利益冲突的商业或财务关系的情况下进行的。
致谢
我们衷心感谢 NCI 生物资源部门提供用于体内研究的 rhIL-2。我们非常感谢提供知情同意以使用皮肤组织进行研究的患者,以及西北大学皮肤生物学和疾病资源中心 (SBDRC) TEST IT 核心的技术援助。
| 


 发表于 2022-6-8 08:47:19
发表于 2022-6-8 08:47:19
 楼主
楼主

